Шейный отдел позвоночника пресмыкающихся представлен
Анатомия, морфология и экология
пресмыкающихся
4. Скелет пресмыкающихся
Позвоночный столб пресмыкающихся разделяется
на пять отделов: шейный, грудной, поясничный,
крестцовый и хвостовой.
Образование гибкой шеи и усиление подвижности
головы имело первостепенное значение при
добывании пищи и при ориентировке. Подвижность
головы обеспечивается дифференцировкой первых
двух шейных позвонков — атласа, или атланта (atlas),
и эпистрофея (epistropheus). Атлас имеет вид костного
кольца, разделенного плотной связкой на верхнюю
и нижнюю половины; через верхнее отверстие
головной мозг соединяется со спинным; передняя
поверхность нижней половины сочленяется с
затылочным мыщелком черепа, а сзади в нижнее
отверстие входит зубовидный отросток второго
шейного позвонка— эпистрофея. Голова может поворачиваться
в стороны на зубовидном отростке, а ее
перемещение в вертикальной плоскости
обеспечивается сочленением черепного мыщелка с
атласом. Все это обеспечивает сложные движения
головы, усиливаемые подвижностью всей шеи.
Изучение эмбрионального развития показало, что
зубовидный отросток образуется путем
прирастания к эпистрофею тела атласа.
Движения шеи обусловлены числом и строением позвонков
шейного отдела; они различны в разных группах. У
гаттерии позвонки еще амфицельные (рыбьего типа)
с остатками хорды между ними. У крокодилов и
большинства чешуйчатых позвонки
процельные (передневогнутые) и лишь у немногих
низших форм амфицельные. Часть шейных позвонков
несет короткие ребра. У скрытошейных черепах,
изгибающих шею в вертикальной плоскости, шейные
позвонки сохраняют лишь зачатки поперечных
отростков. Наоборот, у бокошейных черепах,
изгибающих шею в сторону, поперечные отростки и
связанная с ними мускулатура сильно развиты.
Сложные движения шеи черепах обеспечиваются и
разнообразием позвонков: задние позвонки
процельны, передние — опистоцельны
(задневогнутые), а средний — амфицельный.
К грудным позвонкам причленяются длинные ребра,
брюшные концы которых с помощью хрящевых отделов
прикрепляются к грудине, образуя
замкнутую грудную клетку (грудной
клетки нет у змей). К грудине же
причленяется и плечевой пояс. Поясничные
позвонки тоже несут ребра, которые не доходят до
грудины. К крестцовому отделу, состоящему из двух
позвонков, прикрепляется тазовый пояс.
Хвостовой отдел помогает сохранять равновесие
при передвижении, а иногда служит и движителем (у водных змей, крокодилов и некоторых
водных ящериц).
У ящериц, способных к аутотомии
(отбрасыванию хвоста), хвостовые позвонки могут
разламываться посредине, где имеются тонкие
хрящевые прослойки, делящие тело позвонка на две
части.
Общее число позвонков различно у
разных видов и достигает 50-80 (7-10 шейных, 16-25
грудино-поясничных, 2 крестцовых, 15-40 хвостовых). У
змей и безногих ящериц
число позвонков увеличивается, а позвоночник
делится только на туловищный и хвостовой отделы.
Все туловищные позвонки снабжены подвижными
ребрами, упирающимися в брюшные щитки, что важно
при змееобразном движении. Общее число
позвонков возрастает до 140 (у толстых и коротких
змей) — 435 (у змей с длинным телом).
Парные конечности и их пояса
Плечевой пояс пресмыкающихся сходен с
поясом земноводных, но в нем сильно развиты окостенения.
Коракоид (coracoideum) в месте соединения с лопаткой
(scapula) образуют суставную впадину для причленения
головки плеча (рис. 3).
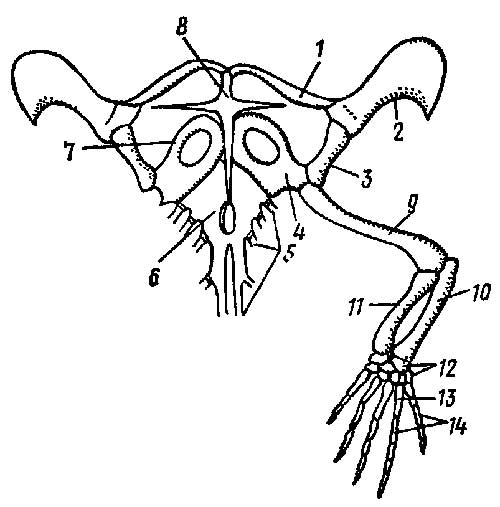
Рис. 3. Плечевой пояс и передняя
конечность ящерицы Lacerta: 1 —
ключица, 2 — надлопаточный хрящ, 3 — лопатка, 4 —
коракоид, 5 — ребра, 6.— грудина, 7 —
прокоракоидный хрящ, 8 — надгрудинник, 9 — плечо,
10 — локтевая кость, 11 — лучевая кость, 12 —
запястье, 13 — пясть, 14 — фаланги пальцев
Сверху к лопатке прирастает уплощенный надлопаточный
хрящ (cartilage suprascapularis), а спереди к коракоиду —
хрящевой прокоракоид (cart. procoracoidea).
Коракоид и прокоракоид каждой стороны
срастаются с непарной костной грудиной
(sternum); через грудную клетку пояс передних
конечностей крепится к осевому скелету. Снизу к
грудине прирастает крестообразная покровная
кость — надгрудинник (episternum). Парные
покровные ключицы (clavicula) соединяют
передний конец надгрудинника с дорзальной
частью каждой лопатки.
Такая конструкция усиливает прочность
плечевого пояса. У черепах
надгрудинник и ключицы входят в брюшной щит
панциря; у крокодилов хорошо развиты лишь
коракоиды и лопатки.
Тазовый пояс состоит из двух безымянных
костей; каждая из них образуется слиянием трех тазовых
костей — подвздошной (ilium), седалищной
(ischium) и лобковой (pubis), совместно
образующих вертлужную впадину, составляющую
сустав с головкой бедра (рис. 14).
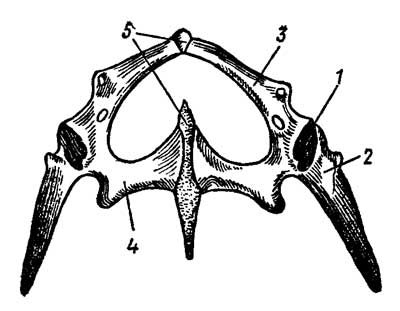
Рис. 4. Тазовый пояс ящерицы
Lacerta: 1 — суставная впадина для головки бедра,
2 — подвздошная, 3 — лобковая, 4 — седалищная
кости, 5 — симфиз
Подвздошные кости сочленяются с поперечными
отростками крестцовых позвонков. У всех
современных пресмыкающихся таз закрытый:
правые и левые лобковые и седалищные кости
соединяются друг с другом по средней линии симфизом
— хрящевой перемычкой.
Парные конечности отличаются у разных
видов и групп пресмыкающихся в зависимости от
преобладания тех или иных способов передвижения.
Но обычно они сохраняют общую схему строения
парных конечностей наземных позвоночных.
В отличие от земноводных у пресмыкающихся в
передней конечности подвижный сустав расположен
между двумя рядами косточек запястья (интеркарпальный
сустав), а в задней конечности — между рядами
косточек предплюсны (интертарзальный
сустав).
Череп пресмыкающихся видоизменялся
главным образом в зависимости от характера
питания и способов добывания пищи. От черепа
амфибий он отличается вытянутыми челюстями,
образующими относительно длинное рыло;
широкий и короткий череп амфибий был необходим
при их ротоглоточном механизме дыхания; в то же
время широкий рот способствовал захвату мелкой
добычи при броске на нее. У пресмыкающихся
дыхание обеспечивается работой грудной клетки,
а захват добычи связан с активным
преследованием, при котором вытянутое рыло имеет
преимущество. Такая форма челюстей давала к тому
же возможность отрывать куски от крупной добычи.
То и другое требовало более мощной жевательной
мускулатуры. С ее развитием и усложнением
органов чувств связаны основные изменения
черепа пресмыкающихся.
Череп почти полностью окостеневает (рис. 5).
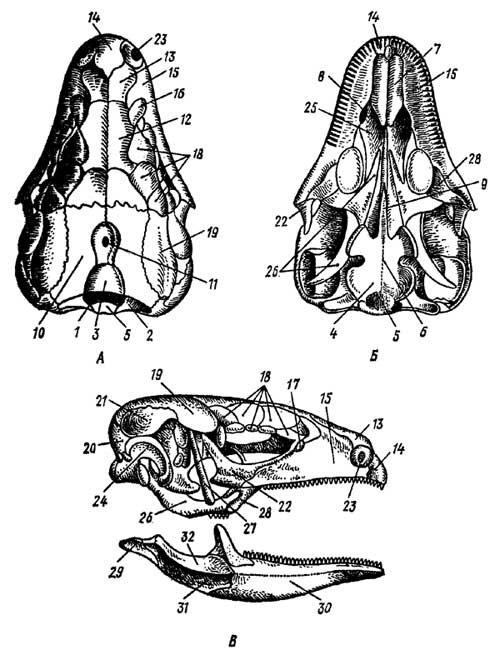
Рис. 5. Череп ящерицы Lacerta
(по Паркеру). А — вид сверху; Б — вид снизу, В — вид
сбоку
1 — затылочное отверстие, 2 — боковая затылочная
кость, 3 — верхняя затылочная кость, 4 — основная
затылочная кость, 5 — затылочный мыщелок, 6 —
основная клиновидная кость, 7 — сошник, 8 — хоана,
9 — парасфеноид, 10 — теменная кость, 11 —
межтеменная кость с отверстием для теменного
органа, 12 — лобная кость, 13 — носовая кость, 14 —
межчелюстная кость, 15 — верхнечелюстная кость, 16
— предлобная кость, 17 — слезная кость, 18 —
надглазничные кости, 19 — заднелобная, или
заглазничная кость, 20 — чешуйчатая кость, 21 —
надвисочная кость, 22 — скуловая кость, 23 —
ноздря, 24 — квадратная кость, 25 — небная кость, 26
— крыловидная кость, 27 — верхнекрыловидная, или
столбчатая кость, 28 — поперечная кость, 29 —
сочленовная кость, 30 — зубная кость, 31 —
надугловая кость, 32 — венечная кость
Затылочная область состоит из четырех затылочных
костей (occipitalia) хондрального происхождения:
верхнезатылочной, основной и двух боковых. Они
окаймляют затылочное отверстие, книзу от
которого лежит единственный затылочный
мыщелок, образованный основной и обеими
боковыми затылочными костями. Покровная
основная клиновидная кость (basisphenoideum)
лежит впереди основной затылочной, образуя дно
черепа. Впереди к ней прирастает небольшой парасфеноид
(parasphenoideum) и располагаются парные сошники
(vomer), сбоку от которых лежат хоаны. В
области слуховой капсулы возникают три ушные
кости (otici); переднеушная, сохраняющая
самостоятельность; заднеушная, срастающаяся с
боковой затылочной, и верхнеушная, срастающаяся
с верхнезатылочной. В обонятельной области
костей нет; она остается хрящевой.
Крыша черепа образована парными покровными
костями: носовыми (nasalia), предлобными
(praefrontalia), лобными (frontalia) и заднелобными
(postfrontalia); далее лежат теменные (parietalia) и
непарная межтеменная (interparietale) кости;
последняя имеет отверстие для теменного органа.
Бока черепа образуют покровные кости:
парные межчелюстные (intermaxillare) у части
видов сливающиеся, парные верхнечелюстные
(maxillare), надглазничные (supraorbitale), скуловые
(jugale), квадратно-скуловые (quadratojugale)
(квадратно-скуловая кость есть в анапсидном и
типичном диапсидном черепе гаттерий и
крокодилов, но исчезает у ящериц, имеющих
диапсидный тип черепа с редуцированной нижней
дугой) и чешуйчатые (squamosum). Небно-квадратный
хрящ в задней части дает начало парным
хондральным окостенениям — квадратным
костям (quadratum), верхним отделом связанным с
мозговой коробкой, а нижним — с нижней челюстью.
Передняя часть нёбно-квадратного хряща
заменяется покровными костями, образующими дно
черепа: парными небными (palatini) и крыловидными
(pterygoidei). Поперечные кости (transversi)
соединяют крыловидные кости с верхнечелюстными,
а у ящериц и гаттерий еще и верхнекрыловидные,
или столбчатые (epipterygoidei), кости связывают
крыловидные кости с теменными.
У черепах и особенно у крокодилов
разрастанием небных отростков межчелюстных и
верхнечелюстных костей, а также небных костей
образуется вторичное костное небо,
разделяющее ротовую полость на верхний отдел — носоглоточный,
и нижний — собственно ротовую полость.
Поэтому хоаны сдвигаются назад, к гортани, что
позволяет дышать, когда из воды выставлен
лишь конец головы с ноздрями.
Нижняя челюсть образуется из меккелева
хряща, замещаемого хондральной костью — сочленовной
(articulare), причленяющейся к квадратной кости, и ряда
кожных костей: зубной (dentale), угловой
(angulare), надугловой (supraangulare), венечной
(coronare) и иногда еще нескольких косточек.
Верхний отдел подъязычной дуги (гиомандибуляре),
как и у земноводных, превращен в слуховую
косточку среднего уха — стремечко (stapes).
Подъязычный аппарат состоит из хрящевой
пластинки (гомолога копулы) и трех пар рожков,
гомологичных гиоидам и остаткам жаберных дуг.
Эволюция жевательной мускулатуры
сопровождалась перестройкой крыши и частично
боковых стенок первичносплошного — стегального
— черепа, имевшегося у земноводных — предков
рептилий. В отдельных ветвях пресмыкающихся
эволюция шла разными путями. В группе анапсида
(ископаемые котилозавры и их непосредственные
потомки — черепахи) первичная стегальная
(сплошная) крыша черепа сохранялась. В ней
образовалась лишь задняя вырезка, освободившая
место для жевательной мускулатуры. В группе
диапсида (гаттерия, крокодилы) образовались две
височные ямы (рис. 6), ограниченные двумя костными
дугами.
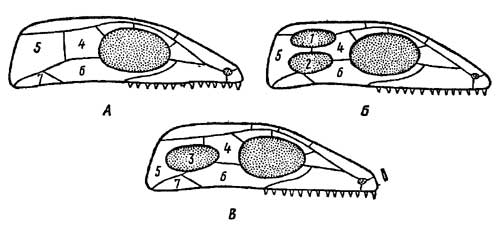
Рис. 6. Схема эволюционных
преобразований стегального черепа у
пресмыкающихся (по Гудричу). А—стегальный
череп; с изменениями свойствен анапсидам; Б —
диапсидный череп с двумя височными ямами; В —
синапсидный череп с одной боковой височной ямой:
1 — верхняя височная яма, 2 — нижняя височная яма,
3 — единственная боковая височная яма, 4 —
заднелобная (заглазничная) кость, 5 — чешуйчатая
кость, 6 — скуловая кость, 7 — квадратно-скуловая
кость
У ящериц диапсидный тип черепа с
редуцированной нижней дугой, а у птиц — с
редуцированной верхней дугой; у змей
обе дуги редуцируются. В группе синапсид — у
звероящеров (тероморфных рептилий) и происшедших
от них млекопитающих — образовалась одна
боковая яма и ограничивающая ее сложная дуга из
элементов верхней и нижней дуг: jugale — quadratojugale —
squamosum. Строение скуловых дуг еще раз подчеркивает
своеобразие группы звероподобных
пресмыкающихся, противопоставляя их другим
группам этого класса.
Параллельно усложнению скелета изменилась структура
костной ткани. Общий тип трубчатых костей
конечностей сохранился, но грубоволокнистая
костная ткань земноводных заместилась тонковолокнистой
костью, обладающей значительно большей
прочностью. При этом кости конечностей приобрели
остеонную структуру и зонарное
(слоистое) строение.
Костный элемент — остеон —
представляет собой костную трубочку, стенки
которой образованы тонкими, но прочными костными
пластинками, а полость занята питающим костную
ткань кровеносным сосудом и нервом. Остеоны в
стенках трубчатой кости располагаются более или
менее концентрически, образуя зонарную
структуру. Внутренняя полость трубчатой кости
занята костным мозгом двух типов: красный
выполняет функцию кроветворной ткани, желтый
участвует в костеобразовании, разрушая кость со
стороны внутренней полости. Обратный процесс —
нарастание кости в толщину — происходит с
наружной поверхности и обеспечивается ее
оболочкой — периостом. Рост трубчатых
костей в длину у рептилий происходит в средней
части трубки (у млекопитающих в местах пере хода
трубки — диафиза в сочленовые головки —эпифизы)
(рис. 7).
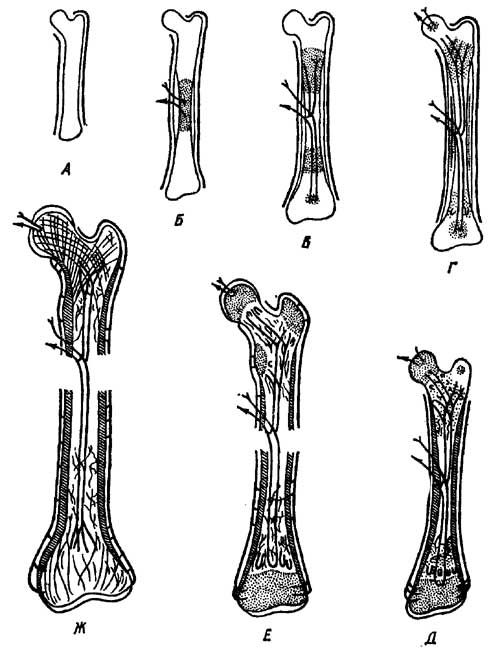
Рис. 7. Развитие трубчатой кости
пресмыкающихся. А — хрящевой зачаток; Б-Ж —
последовательные стадии развития
периостального и эндохоидрального окостенений
(Заварзин и Румянцев, 1946).
Кость такого строения при одинаковом весе и
размерах может выдержать значительно большее
напряжение, нежели кость земноводных. За счет
совершенствования костной ткани, не увеличив
веса скелета, пресмыкающиеся могут выполнять
большую работу. Это увеличило их подвижность.
Уменьшение относительной массы скелета
позволило перейти даже к полету, чему
способствовала пневматизация костей,
внутренние полости которых у птерозавров
частично заполнялись воздухом.
Мускулатура пресмыкающихся, по сравнению с
земноводными, претерпела существенные перестройки.
Исходное метамерное расположение туловищной
мускулатуры, характерное для рыб, в большей части
утеряно уже земноводными. Сложная
дифференциация продолжалась и у рептилий.
Метамерность сохранилась лишь в размещении мышц,
соединяющих рядом расположенные позвонки, и
отчасти в мускулатуре брюшной стенки.
На черепе в височных ямках развивается мощная жевательная
мускулатура, приводящая в движение челюсти.
Появление подвижного шейного отдела
сопровождалось развитием шейной мускулатуры,
особенно мощной у видов с длинной и гибкой шеей.
Лентовидные мышцы туловищной мускулатуры
участвуют в изгибании тела и в движении
конечностей. Непосредственно на последних
размещена тоже мощная мускулатура, особенно
парные сгибатели и разгибатели. Их
совокупная работа обеспечивает разнообразную и
во многих случаях весьма сложную работу
конечностей. Появляются зачатки подкожной
мускулатуры, изменяющей положение роговых
чешуи. Заметно сильнее она развита у змей
и безногих ящериц и участвует в
перемещении тела, обеспечивая координированные
движения брюшных щитков.
В связи с переходом рептилий полностью на легочное
дыхание потребовалось усиление вентиляции
легких. Последнее было обеспечено
возникновением грудной клетки, приводимой в
движение специальной межреберной мускулатурой.
На изменении объема грудной полости сказывается
и движение конечностей (особенно у черепах).
Быстрое передвижение требовало стабилизации
положения внутренних органов. Это было
достигнуто значительным усилением мышц
брюшного пресса.
На нашем сайте Вы также можете познакомиться с информацией по анатомии, морфологии и экологии пресмыкающихся:
общая характеристика пресмыкающихся,
покровы,
движение, и
скелет пресмыкающихся,
органы пищеварения и питание,
органы дыхания и газообмен,
кровеносная система и кровообращение,
органы выделения и водно-солевой обмен,
половые органы и размножение,
нервная система и органы чувств,
поведение и образ жизни,
годовой цикл жизни,
географическое распространение и роль в биоценозах,
значение пресмыкающихся для человека, а также:
некоторые правила зоологической номенклатуры,
определение рептилий по внешним признакам,
рекомендуемая литература по рептилиям.
Познакомиться с изображениями и описаниями других объектов природы России и сопредельных стран —
минералов и горных пород,
почв,
грибов,
водорослей,
лишайников,
листостебельных мхов,
деревьев, кустарников, кустарничков и лиан,
травянистых растений (цветов),
ягод и других дикорастущих сочных плодов,
водных беспозвоночных животных,
насекомых-вредителей леса,
дневных бабочек,
пресноводных и проходных рыб,
земноводных (амфибий),
пресмыкающихся (рептилий),
птиц, птичьих гнезд, их яиц и голосов, а также
млекопитающих (зверей), —
можно в разделе Природа России нашего сайта.
В разделе Природа в фотографиях
размещены также тысячи научных фотографий грибов, лишайников, растений и
животных России и стран бывшего СССР, а в разделе
Природные ландшафты мира — фотографии природы
Европы,
Азии,
Северной и
Южной Америки,
Африки,
Австралии и Новой Зеландии и
Антарктики.
В разделе Методические материалы
Вы также можете познакомиться с описаниями разработанных экологическим центром «Экосистема»
печатных определителей растений средней полосы,
карманных определителей объектов природы средней полосы,
определительных таблиц «Грибы, растения и животные России»,
компьютерных (электронных) определителей природных объектов,
полевых определителей для смартфонов и планшетов,
методических пособий по организации проектной деятельности школьников и полевых экологических исследований
(включая книгу для педагогов «Как организовать полевой экологический практикум»), а также
учебно-методических
фильмов по организации проектной исследовательской деятельности школьников в природе.
Приобрести все эти материалы можно в нашем некоммерческом Интернет-магазине.
Там же можно приобрести mp3-диски Голоса птиц средней полосы России и
Голоса птиц России, ч.1: Европейская часть, Урал, Сибирь.
Источник